The on going evolution of plants.
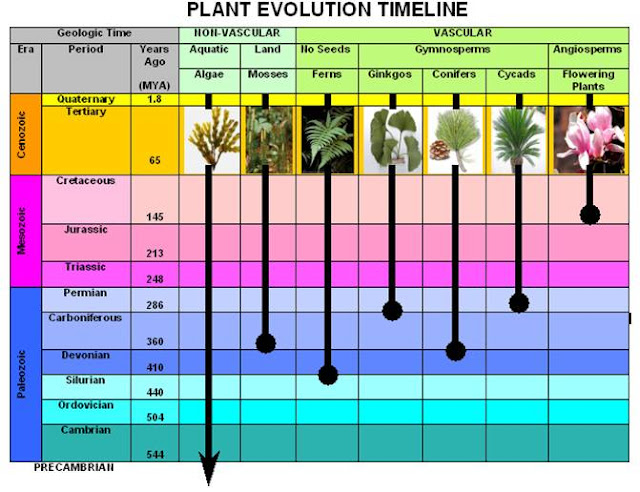
Over the past 10 weeks we have embarked on a journey from 1.2 billion years ago to the present. This final blog is a summary of my previous posts on the evolution of plants. The transition of aquatic plant life to terra-firma during the mid-Paleozoic era enabled the commencement of terrestrial life. Phylogenetic studies linked the ancestral origin of land plants to a charophycean green algae (Crane & Kenrick 1997). During the Upper Ordovician period 475 million years ago, the terrestrialization of charophycean algae led to the evolution of the first permanent land-dwelling plants, the bryophytes (Willis & McElwain 2014). We learnt that bryophyte is used to describe plants with no vascular or specialised transportation system such as liverworts, mosses and hornworts. The evolution of seedless vascular plants towards the end of the Silurian period, 425 million years ago, allowed for the re-transformation of the pre-historic landscape filled with lycophytes (quillw...





